


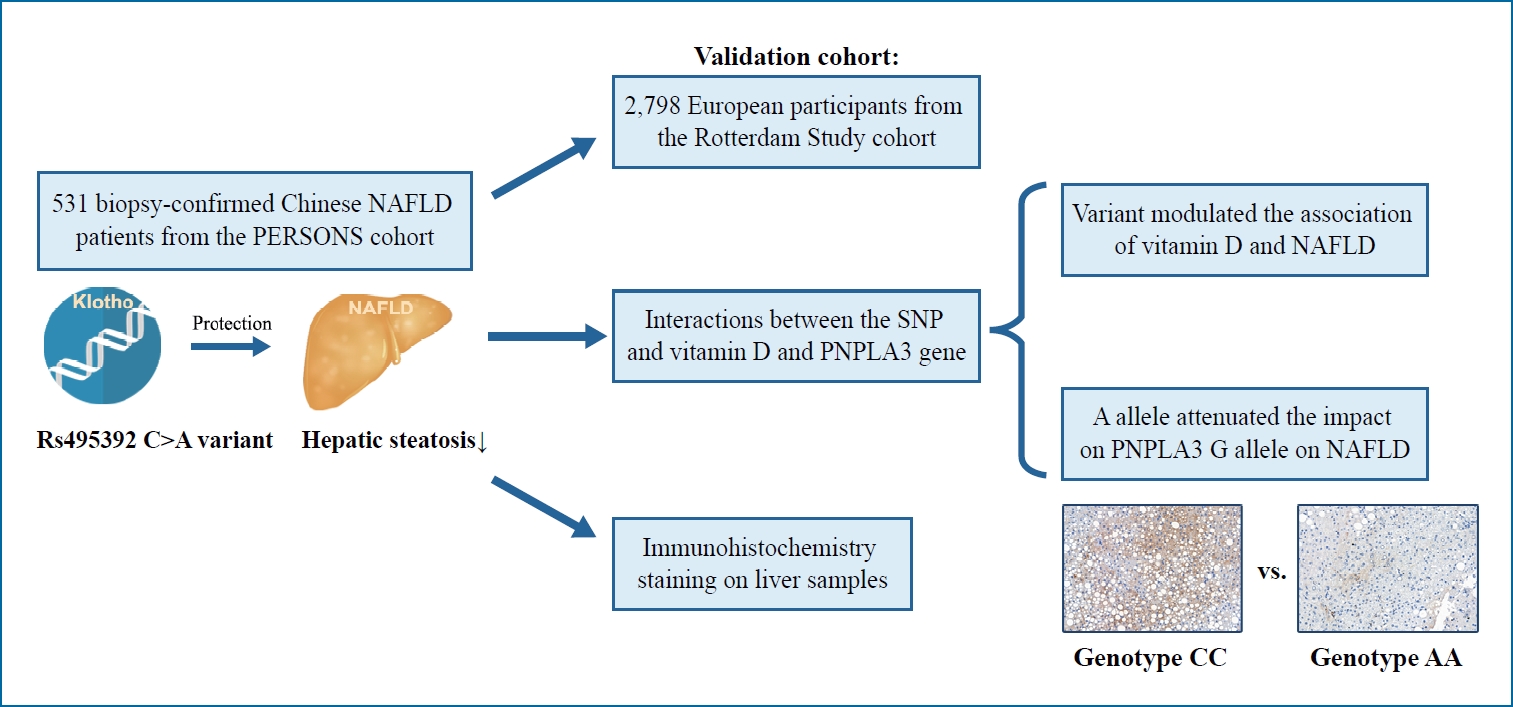
Graphical Abstract
INTRODUCTION
Non-alcoholic fatty liver disease (NAFLD) is shown to be closely associated with metabolic dysfunction including insulin resistance (IR), dyslipidemia, and obesity [1]. NAFLD comprises a spectrum of diseases, ranging from simple steatosis to varying degrees of fibrosis and cirrhosis [2]. In the last decade, more than one-quarter of the global population is suffering from the disease [3]. and an effective pharmacotherapy has yet to be approved. The metabolic dysfunction in fatty liver disease arises from multiple factors, with genes acting as important modifiers. In recent years, multiple large genetic studies have enriched our understanding of the genetic determinants of fatty liver disease [4]. For instance, the patatin-like phospholipase domain containing 3 (PNPLA3) rs738409 C>G variant (p.I148M) has been identified as a major genetic determinant contributing to hepatic fat accumulation and liver fibrosis [4,5]. The potential value of genetic modifiers not only lies in understanding of the pathogenesis of fatty liver disease but also in the designs of new therapeutics.
Klotho, also known as alpha-Klotho, is a well-documented anti-aging protein. It mediates various physiological functions including maintenance of phosphate and calcium homeostasis, attenuation of oxidative stress, suppression of inflammation, and regulation of energy metabolism [6]. It is primarily expressed in the kidney, parathyroid gland, brain, and adipose tissue as a singlepass transmembrane glycoprotein and serves as co-receptor for fibroblast growth factor 23 (FGF23) [7]. There are at least two isoforms of Klotho, a transmembrane form and a soluble form, named soluble Klotho (sKL) in the blood circulation [8]. A previous study using animal models observed that Klotho-knockout mice experienced extreme leanness and a significant reduction of hepatic fat accumulation [9], suggesting a possible direct role in the pathophysiology of fatty liver disease. Analyses of human Klotho (KL) gene polymorphisms revealed their impact on bone density [10], metabolic syndrome [11], chronic kidney disease [12], cardioembolism [13], and coronary artery diseases [14]. Recently, KL rs495392 intronic variant has been suggested to be associated with a decreased risk of atheromatosis and vascular calcification in patients with chronic kidney disease [15,16]. The pathophysiology of atheromatosis is an accumulation and swelling in artery walls made up of macrophage cells, or debris, and contain, among others, lipids (cholesterol and fatty acids). The single nucleotide polymorphism (SNP) rs495392 locates in an intronic region of the KL gene; thus, no variations in the Klotho protein sequence are expected. However, rs495392 has been shown to affect the KL gene expression in blood in previous expression quantitative trait loci (eQTL) data [17]. Given the possibility of altering KL expression and the role in lipid accumulation, the KL rs495392 polymorphism may have an impact on the susceptibility to fatty liver disease. Therefore, it is pivotal to explore the effect of rs495392 in liver disease.
Vitamin D exerts diverse physiological effects on calcium and phosphate homeostasis, insulin sensitivity, anti-inflammation, and anti-fibrosis [18]. Some studies have suggested that serum vitamin D status was associated with the severity of fatty liver disease and its supplementation could improve insulin sensitivity in patients with fatty liver disease [19-21]. It has been shown that active vitamin D can upregulate the expression of the KL gene through vitamin D-responsive elements, while Klotho/FGF23 axis suppresses the activation of vitamin D [22]. Therefore, it is also important to speculate whether KL rs495392 polymorphism alters the effect of vitamin D on fatty liver disease.
The aims of this study were: 1) to explore the association of KL rs495392 polymorphism with hepatic steatosis in patients with NAFLD, 2) to clarify whether the role of vitamin D on NAFLD is different under the KL gene variant, and 3) to discern if there is any interaction between the PNPLA3 rs738409 and KL rs495392 gene polymorphisms.
MATERIALS AND METHODS
Design and study population
The study population was individuals recruited from the PERSONS project (January 2017 to September 2020). The diagnostic criteria for NAFLD have been described previously [23]. Briefly, patients with biopsy-confirmed fatty liver without a secondary liver disease (including alcohol use disorder, viral hepatitis, autoimmune hepatitis, hepatocellular carcinoma, etc.) have been included. A total of 531 Chinese patients with biopsy-confirmed NAFLD were enrolled.
Clinical data and laboratory examinations
Medical history, former drug use information and other clinical information is recorded at the baseline. Biochemical data were collected from all patients within 24 hours of liver biopsy. Body mass index (BMI) was calculated as weight (kg) divided by the square of height (m). IR was estimated according by the homoeostasis model assessment (commonly referred to as HOMA) [24]. The threshold for IR was defined as HOMA-IR (homeostasis model assessment-insulin resistance) score ≥2.73 [25]. Diagnosis of diabetes was based on criteria of the American Diabetes Association [26]. Serum level of 25-hydroxyvitamin D [25(OH)D] was measured to show vitamin D status and defined as: vitamin D sufficiency ≥75 nmol/L, insufficiency 50–75 nmol/L, and deficiency <50 nmol/L.
Histological assessment of the liver
Liver biopsies were performed using a 16-gauge needle under ultrasound guidance. The histology was reviewed by a single liver pathologist (X.D.W.) who was blinded to the clinical and biochemical data. Histological parameters were defined based on the NAFLD Activity Score (NAS) system by Kleiner et al. [27]. Severe steatosis, severe ballooning, severe lobular inflammation, and significant fibrosis were defined if their scores were ≥2.
Immunohistochemistry (IHC) staining
IHC staining was carried out on liver samples to observe the presence of Klotho protein. First, the sections were treated with a citric acid antigen retrieval buffer for antigen retrieval. After blocking endogenous peroxidase activity, sections were incubated with primary antibodies in a washing buffer for 12 hours at 4°C Then, tissue slides were washed thrice with phosphate buffered saline, and an appropriate secondary antibody was added. Finally, the sections were treated with streptavidin-horseradish peroxidase complex and diaminobenzidine solution, and counterstained with hematoxylin. The primary antibody was against sKL (Abcam, Cambridge, MA, USA). Images of the sections were acquired by light microscopy, using a Adimec Q-12A-180Fc microscope (Adimec, Eindhoven, the Netherlands). The semiquantitative analysis of Klotho expression was measured by Image Pro-Plus v. 6.0 software (Media Cybernetics, Bethesda, MD, USA).
SNP genotyping
The genotyping of KL rs495392 C>A and PNPLA3 rs738409 C>G variants was performed using the MassARRAY (Agena Biosciences, San Diego, CA, USA) according to the manufacturer’s protocol. For the purpose of genotyping, each sample used approximately 20 ng of genomic DNA. The primer sequences were the following: forward 5’–ACGTTGGATGGGAGGAAGAAATACTTGAGAC–3’, reverse 5’–ACGTTGGATGTAATTGTAACACCCTTGTGC–3’ (rs495392); forward 5’–ACGTTGGATGTCTCCTTTGCTTTCACAGGC–3’, reverse 5’–ACGTTGGATGCTGAAGGAAGGAGGGA TAAG–3’ (rs738409).
Statistical analysis
Statistical analyses were performed using R software (v3.5.2; R Foundation for Statistical Computing, Vienna, Austria), SPSS (ver. 20.0; SPSS Inc., Armonk, NY, USA). Continuous variables are expressed as means±standard deviations for those with a normal distribution and as medians (p25–p75) for those with a non-normal distribution. Categorical variables are expressed as frequency (%). Comparisons between groups were done using t test, Mann-Whitney test, and chi-squared test, respectively. The Hardy-Weinberg equilibrium (HWE) was assessed using the one tailed chi-square test. Multivariate logistic regression models were performed to test the association between the SNP and liver histology features. Due to the relatively low frequency of the minor allele, the impact of rs495392 on histological damage was modelled under a dominant inheritance model. The SNP-SNP interactions were analyzed by generalized multifactor dimensionality reduction (GMDR). A post-hoc power analysis for the logistic regression models were undertaken to assess the statistical power in present study. A sample size (N) of 531 achieves 73.4% of power (1-β) for KL rs495392 variant and hepatic steatosis analysis. A P-value <0.05 was considered to be statistically significant.
The replication cohort from the Rotterdam Study (RS)
The RS was used as replication cohort. The RS is a prospective cohort study of individuals aged ≥40 years living in the Ommoord district of Rotterdam, the Netherlands. The objectives, design and methods of the RS have been described in detail elsewhere [28]. In 1990, the first cohort of study (RS-I) comprised 7,983 persons aged ≥55 years. In 2000, the second cohort (RS-II) was extended to include an additional 3,011 participants who moved into the study district or had become 55 years of age. A further extension of the cohort (RS-III) formed in 2006 and include 3,932 participants living in the research area and aged ≥45 years. In 2016, the recruitment of another extension (RS-IV) started that targeted participants aged ≥40 years. All participants in the study provided written inform consent to participate and to obtain information from their treating physicians.
Genotyping was performed using Human Core-24 BeadChip Marker information, and the quality control procedures were applied to the genotype data. Data were imputed using the second release of the Haplotype Reference Consortium (realizes 1.1 2016) by mean Michigan Imputation Server. We extracted genotype dosage for rs495392 with a high imputation quality (Rsq score=0.98).
For the current study, 4,307 participants from the RS-I, RS-II, and RS-III sub-cohorts who had information on the KL rs495392 polymorphism and fatty liver index (FLI) were included. Then, the participants with excessive alcohol consumption, medicine use that cause steatosis, other viral hepatitis or data missing were excluded, leaving 3,830 participants included as the main population for analysis (Supplementary Fig. 1). The definitions of NAFLD and fibrosis for the participants were based on FLI and liver stiffness measurement (LSM), respectively. According to the previous studies, a FLI ≥60 was consider as hepatic steatosis [29], while a LSM >6.9 kPa was consider as significant fibrosis [30].
Ethics
Ethical approval for the PERSONS project was obtained from the Ethics Committee of the First Affiliated Hospital of Wenzhou Medical University (2016-246, 1 December 2016). All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional research committee and with the 1964 Helsinki declaration and its later amendments.
The RS has been approved by the Medical Ethics Committee of the Erasmus MC (registration number MEC 02.1015) and by the Dutch Ministry of Health, Welfare and Sport (Population Screening Act WBO, license number 1071272-159521-PG).
Written informed consent was obtained from all participants included in the study.
RESULTS
Genotype distribution
A total of 531 genotyped patients (from the PERSONS project) were included in this study. The frequency distribution of KL rs495392 was in HWE (P>0.05). The minor allele frequency for KL rs495392 was 0.168 in our Chinese cohort, similar to that (0.188) reported for the general East Asian population of the 1000 genomes project [31]. The genotype distribution of rs495392 CC, CA, and AA was 68.9%, 28.6%, and 2.4%, respectively.
Patient characteristics
The characteristics of patients with biopsy-confirmed NAFLD stratified by carriage of KL rs495392 A allele are depicted in Table 1. The comparison of the characteristics according to the rs495392 genotypes showed that the group with genotype CA+AA had older average age, lower serum level of aspartate aminotransferase, and higher serum levels of high-density lipoprotein cholesterol (HDL-C) than the group with CC genotype (all, P<0.05).
Impact of KL rs495392 polymorphism on the liver histology features
The impact of carriage of KL rs495392 A allele on the liver histology features is presented in Table 1. The carriers of the A allele (CA+AA) had significantly lower hepatic steatosis score, NAS and prevalence of severe steatosis, compared to the C allele carriers (all, P<0.05).
To further estimate the effects of rs495392 on histological damage, logistic regression models were performed in Table 2. In crude analysis, carriage of the A allele was related to a lower risk of severe hepatic steatosis with an odds ratio (OR) of 0.61 (CA+AA vs. CC, 95% confidence interval [CI], 0.42–0.89; P=0.010). After adjusting for age, sex, BMI, and presence of diabetes, the A allele still showed a protective effect against severe steatosis with an OR of 0.66 (CA+AA vs. CC; 95% CI, 0.45–0.98; P=0.040). However, there was no significant association between rs495392 and hepatocyte ballooning, lobular inflammation, and fibrosis.
Hepatic sKL protein expression
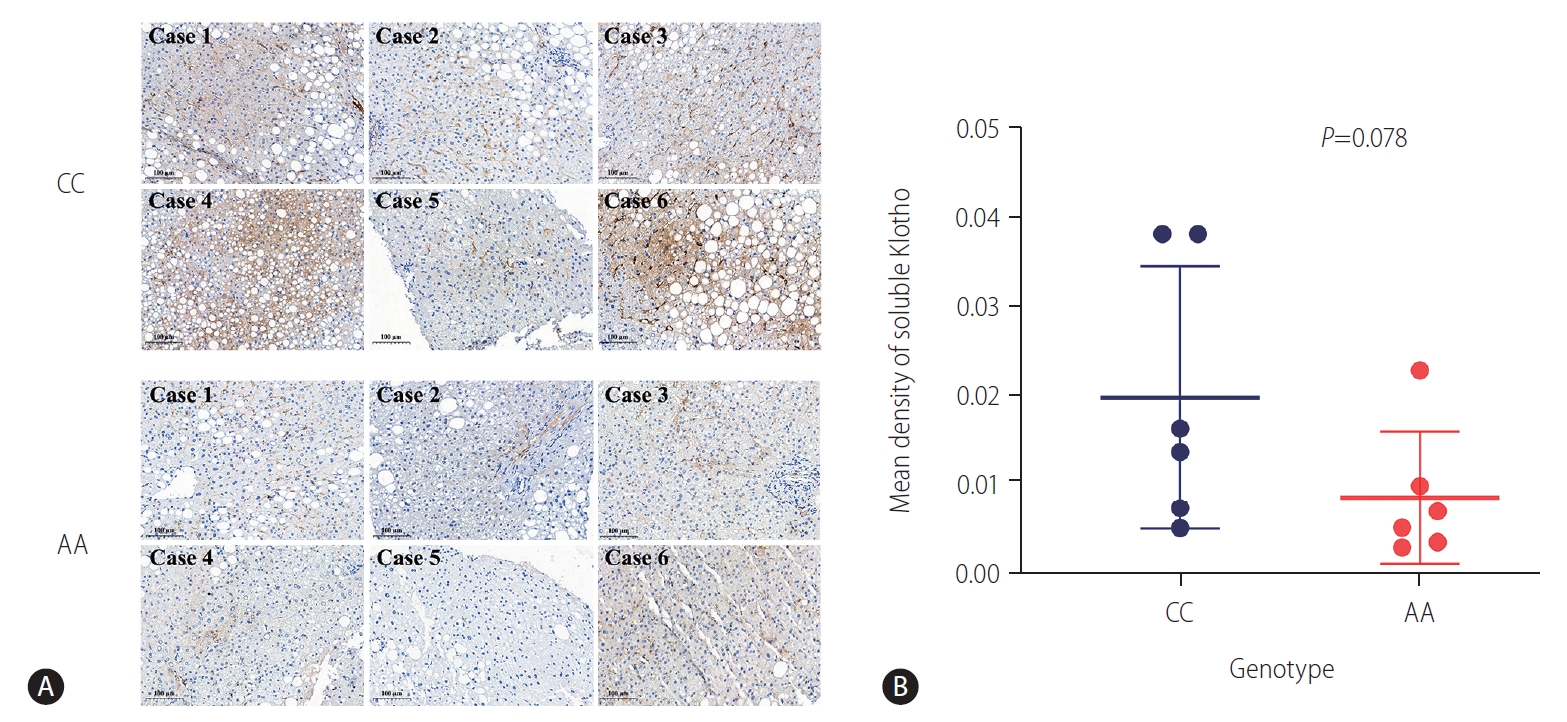
To determine whether KL protein is expressed differentially in liver according to the genotypes, 12 homozygous liver samples underwent IHC staining (six from the genotype CC patients and six from the genotype AA patients with matched age, sex and presence of diabetes). According to the instructions of KL antibody, only soluble form of the protein can be detected. As shown in Figure 1A, a reduced expression of sKL seemed to present in the genotype AA group compared with the CC group. After quantifying sKL expression, we found that the liver samples from the genotype AA showed a non, but borderline, significant lower level of sKL (Fig. 1B; P=0.078). Considering the limited sample size and low statistical power, it is possible that the KL rs495392 polymorphism may downregulate the sKL expression.
Replication of the impact of KL rs495392 polymorphism on hepatic steatosis in the RS
To validate the effect of KL rs495392 polymorphism in an external cohort, we obtained data from the prospective, population-based RS cohort. The characteristics of participants from the RS are shown in Supplementary Table 1. As shown in Supplementary Figure 1, a total of 1,143 NAFLD and 2,687 non-NAFLD participants of the European ancestry were included. The median value of FLI was 78 in the NAFLD participants. As demonstrated in Table 3, the genotype AA was negatively associated with higher FLI (OR, 0.36; 95% CI, 0.17–0.73; P=0.006), indicating a protective effect of the A allele against having higher FLI score. After stratifying by severity of liver fibrosis, the protective effect of A allele was only observed in participants with less fibrosis (OR, 0.17; 95% CI, 0.05–0.49; P=0.002). In addition, the KL rs495392 A allele was also associated with a reduced risk of type 2 diabetes in this Caucasian cohort (Supplementary Table 1).
The role of vitamin D in NAFLD under the KL rs495392 polymorphism
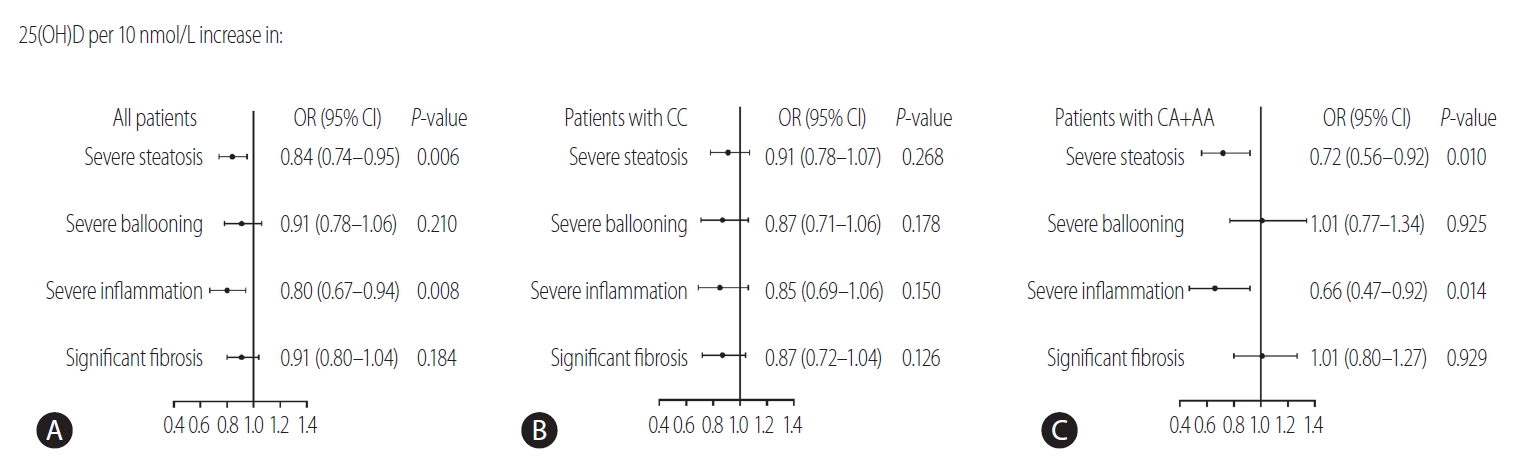
To investigate whether KL rs495392 polymorphism influences the association of vitamin D with NAFLD, the serum levels of 25(OH)D were measured according to the rs495392 genotype in our Chinese patient cohort. As shown in Table 1, the patients carrying the A allele (CA+AA) had significant higher serum levels of 25(OH)D, compared to the patients with the CC genotype (58.0±19.1 vs. 63.5±19.8 nmol/L, P=0.032, respectively). Furthermore, patients were classified into three groups according to the serum 25(OH)D levels (deficiency [<50 nmol/L] vs. insufficiency [50–75 nmol/L] vs. sufficiency [≥75 nmol/L]). The prevalence of severe steatosis and severe inflammation was stepwise deceased from deficient to sufficient status of vitamin D (all P<0.05; P for trend <0.05) (Supplementary Fig. 2). Our results suggested that a serum level of 25(OH)D could be negatively associated with the severity of NAFLD. After stratification for the rs495392 genotypes, the association between serum level of 25(OH)D and liver damage was only evident in patients who were carrying the A allele, CA or AA genotype (OR, 0.72; 95% CI, 0.56–0.92; P=0.010 for severe steatosis and OR, 0.66; 95% CI, 0.47–0.92; P=0.014 for severe inflammation) (Fig. 2).
Interaction between PNPLA3 rs738409 and KL rs495392 polymorphisms
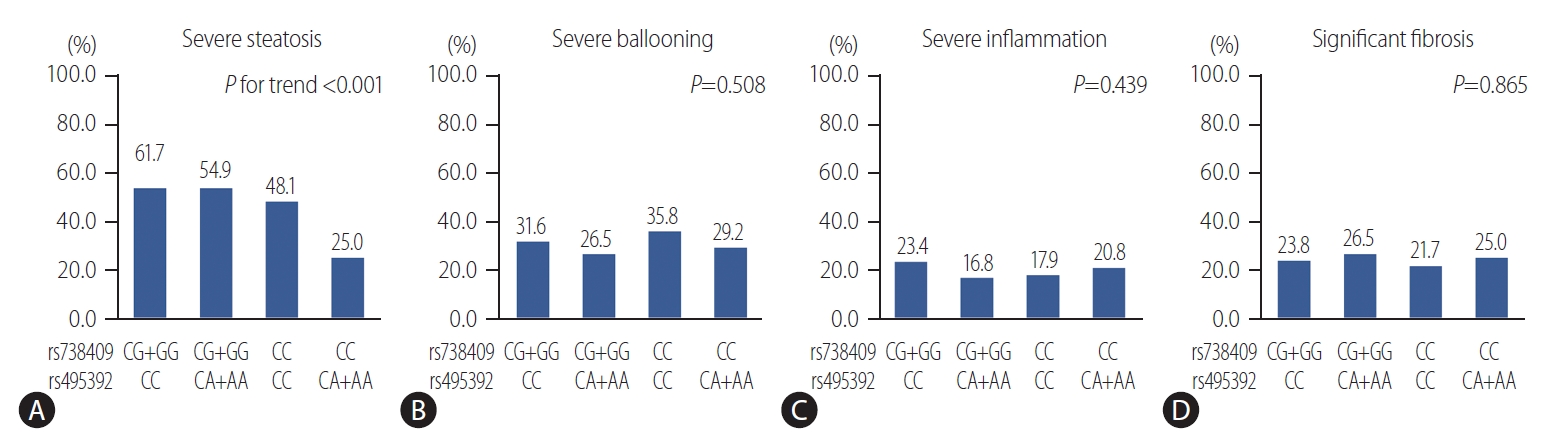
The PNPLA3 rs738409 polymorphism is known as a main genetic determinant of fatty liver disease [5]. Therefore, we next explored the interaction between PNPLA3 rs738409 and KL rs495392 variants on the liver damage. The results of the GMDR model analysis of the SNP-SNP interactions are showed in Table 4. The best model for predicting severe steatosis was the two-locus model, a combination of PNPLA3 rs738409 and KL rs495392, with the highest testing accuracy (TA) and perfect cross-validation consistency (CVC) (TA, 59.1%; P=0.001; CVC, 10/10). The prevalence of severe steatosis according to the rs738409/rs495392 genotypes were 61.7% for CG+GG/CC, 54.9% for CG+GG/CA+AA, 48.1% for CC/CC, 25.0% for CC/CA+AA (P for trend <0.001) (Fig. 3). The prevalence of severe steatosis stepwise changed according to the comprehensive effects of rs738409 and rs495392 polymorphism. To further investigate these interactions, logistic regression models were used according to rs738409 and rs495392 genotypes (Table 5). Using patients with wildtype genotype as reference, patients with the rs495392 A allele had a lower risk of severe steatosis with an OR of 0.38 (95% CI, 0.17–0.84; P=0.016), while patients with rs738409 G allele had an increased risk of severe steatosis with an OR of 1.68 (95% CI, 1.04–2.72; P=0.035). Patients with both alleles of rs495392 A allele and rs738409 G allele showed no significant difference compared with the wildtype alleles (OR, 1.39; 95% CI, 0.79–2.44; P=0.249). Thus, it might be possible that the KL rs495392 A allele attenuates the detrimental impact of the PNPLA3 rs738409 G variant on hepatic fat accumulation in patients with NAFLD. However, no interactions between rs738409 and rs495392 were observed on other histology features of NAFLD (Table 5).
DISCUSSION
In this study, we examined the impact of a polymorphism at the KL locus, namely the rs495392 C>A intronic variant previously associated with cardiometabolic traits [15,16], on liver damage in a cohort of Chinese patients with biopsy-confirmed NAFLD and an external European population-based cohort, the RS. Our results suggest that the rs495392 A allele has a protective impact on the severity of hepatic steatosis. In further analysis, rs495392 variant modulated the association of vitamin D with hepatic steatosis and lobular inflammation in NAFLD. Furthermore, we also found that the rs495392 A allele may attenuate the detrimental impact of PNPLA3 rs738409 G allele, one of the main genetic determinants of fatty liver disease [4], on the risk of severe steatosis.
Recent studies have identified a role of Klotho, known as an antiaging gene, in energy metabolism [6,32,33] and observed a therapeutic potential in cell and animal models with metabolic disorders [34,35]. In vitro studies, Klotho has been detected in adipogenic cell and identified as an adipogenesis promoting factor, which promote adipocyte differentiation [36]. In parallel with the findings, the Klotho-knockout mice experienced extreme leaness. Even in leptin-deficient mice, eliminating Klotho function resulted in significant reduction of hepatic fat accumulation and the whole body weight [9]. Inhibition of Klotho expression presented inspiring benefit in improving lipid accumulation and obesity. However, the physiological effects of Klotho in metabolism are complicated and controversial. Gu et al. [37] observed that the increased serum lipid profiles and hepatic lipid content in sKL deficient type 2 diabetes mice. Nevertheless, the overexpression of sKL reversed these phenomena [37]. Rao et al. [35] found administration of Klotho resulted in decreased fat mass, increased lean mass and reduced lipid accumulation in liver of obese mice. In addition to hepatic lipid accumulation, IR also plays a crucial role in fatty liver disease [38,39]. Interestingly, Klotho overexpressing mice experience IR but without hyperglycemia and obesity. Mice with deficient Klotho gene expression showed increased insulin sensitivity, and experienced many aging-associated disease phenotypes [40,41]. Despite the prominent roles of Klotho in peripheral metabolism, studies demonstrated a central function of Klotho in cerebrospinal fluid (CSF). The CSF Klotho regulates energy balance and intracerebroventricular administration of Klotho suppressed food intake, improved glucose profiles, and reduced body weight in mice [42,43]. The molecular mechanisms of Klotho have not been fully elucidated so far, while peroxisome proliferator-activated receptor (PPAR) family highly involves in Klotho gene expression. PPAR family is recognized as an important regulator of energy metabolism over the past decades [44]. The Klotho gene contains a non-canonical PPAR-response element in its 5ʹ-flanking region, and the binding of PPAR-γ to this element induces Klotho gene expression [45]. Moreover, PPAR-α was also reported to mediate the regulation of sKL on hepatic lipid metabolism in mice [37].
Genetic polymorphisms of Klotho gene were associated with various pathophysiological processes and diseases [10-13]. In our Chinese cohort, carriage of the KL rs495392 A allele was related to a lower risk of severe steatosis in patients with NAFLD, without affecting body mass nor IR. In replication cohort, the RS from the Netherlands, we observed that rs495392 A allele was not only associated with lower FLI, but also a reduced risk of type 2 diabetes (Supplementary Table 1), indicating the different effects of the variant between Chinese and European populations. Rs495392 locates in an intronic region of the KL gene. Notably, intronic variants have been reported to affect gene expression and play a role in, for example, alternative splicing of the mRNA [46]. For instance, another intronic variant in KL rs650439 was suggested to be associated with plasma Klotho concentration [47]. In addition, previous large eQTL meta-analysis in whole blood suggested the SNP rs495392 as cis-eQTL signal for the expression of KL gene [17]. Although the present study does not have the information on serum Klotho concentrations, the expression of sKL in liver tissues was examined. The protection of KL rs495392 variant against hepatic steatosis seemed to be related to a downregulation of sKL expression. Nevertheless, the statistical power of the results is low and further investigation is required.
Moreover, we conducted the analysis of interaction between PNPLA3 rs738409 and KL rs495392 variants. PNPLA3 is a 481-residue protein and mainly expressed in the liver. It has lipase activity towards triglycerides in hepatocytes and retinyl esters in hepatic stellate cells. The PNPLA3 gene is associated with hepatic steatosis and regarded as the main genetic factor for fatty liver disease [4]. In this work, we noted that KL rs495392 A allele may attenuate the risk effect of PNPLA3 rs738409 G allele on hepatic steatosis. Based on current data, the underlying mechanism of the interaction between rs495392 and rs738409 remains unclear, and therefore further experimental studies are needed.
The interactions between Klotho and Vitamin D are complicated. Briefly, Klotho-FGF23 axis can suppress synthesis of 1,25-dihydroxyvitamin D [1,25(OH)2D] from 25(OH)D, while 1,25(OH)2D positively regulates KL gene expression [48]. A recent study has reviewed the efficacy of serum vitamin D and its supplementation in fatty liver disease and suggested inconclusive conclusions [19]. Four cross-sectional studies from Italian, Chinese, and Korean showed the significant relationships between vitamin D and severity of fatty liver disease [49-52]. However, two meta-analyses concluded that there was no association between vitamin D level and fatty liver disease [53,54]. In terms of vitamin D supplementation, animal and cell experiments showed benefits in improving NAFLD-like symptoms, whereas most of the clinical trials failed in treatment of patients with fatty liver disease [55]. In our cohort, the KL rs495392 variant increased serum vitamin D expression. Impaired vitamin D status was associated with advanced hepatic steatosis and lobular inflammation and these effects were inconsistent according to the KL rs495392 genotypes. Hence, the inclusive conclusions of vitamin D in fatty liver disease may be partly attributed to the differential impact of vitamin D status in patients with different genetic background.
The strengths of our study are large sample size, two different population cohorts. Replication in external cohort from different ancestries allowed us to improve the robustness of the findings. However, our study has some limitations. The present work lacks information of serum sKL concentration. Although the IHC staining of liver tissues underwent, the sample size was limited and only non-significant trend was observed. Furthermore, the replication cohort from the RS lacks histology data and the methodology used for assessing NAFLD is not the same as the cohort from China. However, it would be too difficult to use liver biopsy as a diagnostic method for participants of epidemiological studies, and at the population level. Moreover, the form of vitamin D measured in this study is 25(OH)D rather than 1,25(OH)2D. The inactive form of vitamin D is more stable in circulation, but indirectly represents the active form of vitamin D.
In conclusion, this study indicates that the KL rs495392 polymorphism had a protective effect against severe hepatic steatosis in patients with NAFLD. These data suggest that Klotho may represent a therapeutic target for the fatty liver disease that warrant further investigation.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement1
Supplement1 Print
Print





