Eating, diet, and nutrition for the treatment of non-alcoholic fatty liver disease
Article information

Abstract
Nutrition and dietary interventions are a central component in the pathophysiology, but also a cornerstone in the management of patients with non-alcoholic fatty liver disease (NAFLD). Summarizing our rapidly advancing understanding of how our diet influences our metabolism and focusing on specific effects on the liver, we provide a comprehensive overview of dietary concepts to counteract the increasing burden of NAFLD. Specifically, we emphasize the importance of dietary calorie restriction independently of the macronutrient composition together with adherence to a Mediterranean diet low in added fructose and processed meat that seems to exert favorable effects beyond calorie restriction. Also, we discuss intermittent fasting as a type of diet specifically tailored to decrease liver fat content and increase ketogenesis, awaiting future study results in NAFLD. Finally, personalized dietary recommendations could be powerful tools to increase the effectiveness of dietary interventions in patients with NAFLD considering the genetic background and the microbiome, among others.
INTRODUCTION
Non-alcoholic fatty liver disease (NAFLD) is the fastest-growing and most prevalent liver disease worldwide, contributing essentially to liver-related morbidity and mortality [1]. Being a prototype of so-called “non-communicable diseases”, the increasing prevalence of NAFLD, but also obesity, is regarded as closely related to changes associated with modern-day lifestyle including increased calorie intake, reduced physical activity, and sedentary behavior [2] that result in a mismatch between a decreased energy expenditure and an increased energy intake [3,4]. Among other factors [4], this seems to be largely driven by socioeconomic factors leading to a rise in ubiquitous, cheap, and energy-dense food of low dietary quality. In the absence of approved pharmacological treatments, lifestyle and especially dietary interventions are even more important to counteract the growing burden of NAFLD [5]. Here, we provide a concise overview of different nutritional strategies in NAFLD, especially in overweight and obese patients (Fig. 1), and summarize our current understanding of the interplay between NAFLD and our diet to facilitate personalized nutritional advice in these patients.
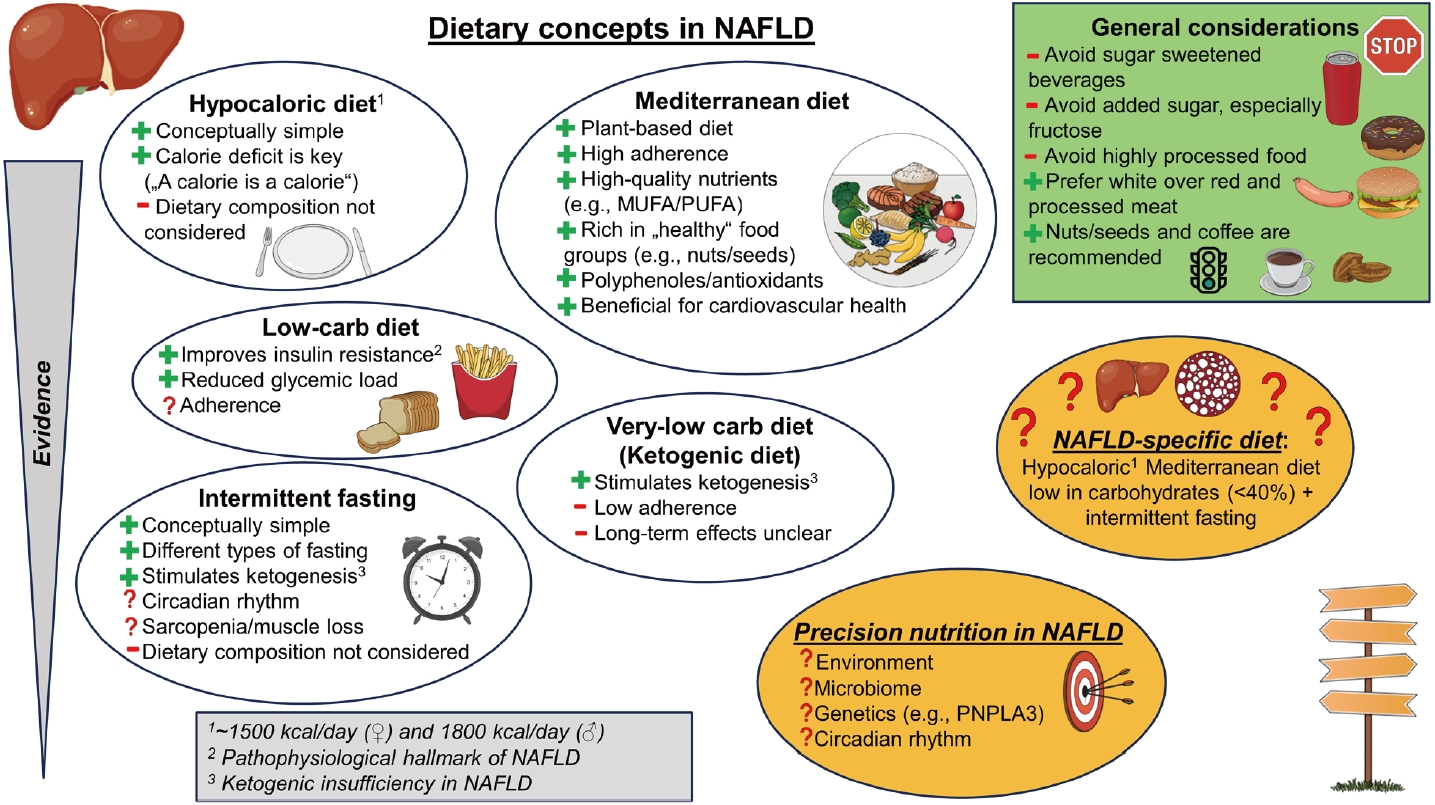
Overview of dietary concepts in NAFLD highlighting evidence, pathophyisological considerations and open questions. NAFLD, nonalcoholic fatty liver disease; MUFA, mono-unsaturated fatty acids; PUFA, poly-unsaturated fatty acids.
Current guideline recommendations
In brief, current European [6,7], American [8], Asian [9], and Korean [10] guidelines highlight the importance of two essential concepts to treat NAFLD in overweight and obese individuals: (I) Weight loss aiming at a reduction of 7–10% in body weight, and (II) energy restriction aiming at a calorie deficit of approximately 500–1,000 kcal/day. On top of these established recommendations, the ideal macronutrient composition is currently a matter of debate: While the American society highlights uncertainties regarding long-term (histological) endpoints that preclude recommendations in favor of one type over another, a dietary composition in accordance to the Mediterranean dietary (MD) is generally advised by European and Asian societies [6,7,9] given clear signals towards beneficial effects beyond the macronutrient composition (see chapter Mediterranean Diet [MD]). Also, the latter advise avoiding added fructose, mostly via consumption of sugar-sweetened beverages (SSB). Importantly, both weight loss and a calorie deficit might be only achieved in combination with an increase in physical activity and exercise that ultimately lead to an increased energy expenditure [5]. Thus, a combined “lifestyle”-approach should always be preferred, and tailored to the individual patient to increase long-term adherence achieving a durable improvement in energy metabolism (“eat less, move more”) [6,8].
Outcomes in nutritional research
To make use of dietary recommendations in clinical practice, one must take the endpoints that have been investigated in the respective studies into account. With this regard, dietary recommendations for NAFLD are especially complex given the variety of clinical endpoints: (I) Improvement of liver histology including regression of fibrosis or resolution of non-alcoholic steatohepatitis (NASH) [11-13]; (II) changes in quantitative parameters assessing liver fat content (i.e., hepatic steatosis) such as the intrahepatic triglyceride content/ intrahepatic lipid content (IHLC) assessed via magnetic resonance spectroscopy [14,15], controlled attenuation parameter (CAP) assessed by transient elastography [16,17], or scores combining laboratory values such as the fatty-liver-index [18,19]; (III) quantitative assessment of liver fibrosis using magnetic resonance elastography [20] or transient elastography-based liver stiffness measurement (LSM) [16-18,21]; (IV) transaminases (aspartate aminotransferase [AST]/alanine transaminase [ALT]) as a surrogate for hepatic inflammation [22]; and (V) changes in metabolic parameters such as fasting blood glucose, insulin resistance, serum lipids but also body weight that do not specifically address changes in the liver. Especially regarding liver fat content, one has to consider its transiency and that presumed association with clinical endpoints are predominantly driven by hepatic fibrosis (e.g., cardiovascular diseases [23]) including mortality [24]. Also, combined scores such as the fatty-liver-index have not been developed for metric assessment of liver fat, making absolute changes in these scores uninterpretable [25]. At the same time, levels of ALT/AST have numerously been described as inadequate to portray disease severity and hepatic fibrosis in NAFLD [26-28]. Finally, studies using histological data are scarce [11-13]. While they would be urgently needed, they are reasonably limited given the invasiveness of liver biopsy. With this regard, trials focusing on accepted surrogates of hepatic fibrosis (such as magnetic resonance elastography or LSM) should be strongly encouraged in future nutritional intervention studies.
CALORIE RESTRICTION & HYPOCALORIC DIET
Clear evidence suggests that dietary calorie restriction is able to improve numerous metabolic parameters beyond its effect on liver-related outcomes (e.g., reviewed in [29]). Focusing on NAFLD, several studies have shown that a total energy deficit (~500 kcal/day resulting in ~1,500 kcal/day for women and ~1,800 kcal/day for men) leads to a decrease in body weight, transaminase levels, total body fat, visceral fat, and IHLC, regardless of how it is achieved [15,22,30,31]. An important study by Kirk et al. [32] (2009) reported similar changes in body weight, body composition, and IHLC after 7% of weight loss (i.e., after around 11 weeks) following a hypocaloric low-carbohydrate diet (LCD) vs. a high-carbohydrate diet (HCD) despite short-term effects in favor of LCD (i.e., after 48 hours) [32]. Again, studies associating the degree of weight loss with the extent of histological improvement [11] and improvement of metabolic parameters [33] strongly favor a dose-dependent effect of nutritional/lifestyle interventions beyond macronutrient composition [34]. Interestingly, a recent meta-analysis of observational studies including >100,000 individuals has shown that the only difference between NAFLD and controls was a higher calorie intake while the macronutrient composition did not significantly differ [35]. Finally, evidence highlighting the importance of calorie reduction originates from the observation that LCD (as discussed in chapter Low-Carbohydrate Diet [LCD]) are only successful in reducing IHLC when integrated into a hypocaloric diet approach, but fail to decrease or even increase IHLC if carbohydrate restriction occurs at the expense of increased fat intake in an isocaloric manner [36,37].
LOW-CARBOHYDRATE DIET (LCD)
On top of calorie restriction, increasing evidence suggests a diet low in carbohydrates to be especially fruitful for patients with NAFLD. On a population-based level, data from America show that intake of potato chips, potatoes, and SSB were the dominant factors associated with weight gain [38] paralleling the global increase in obesity and NAFLD in recent years, thereby clearly suggesting a certain role of a western diet typically high in carbohydrates for the surge in obesity and NAFLD. On the short term, Browning et al. [39] (2011) reported a favorable reduction in IHLC after a hypocaloric LCD (8% carbohydrates [C], 33% protein [P], 59% fat [F]) compared to a hypocaloric diet (50% C, 16% P, 34% F), as did Kirk et al. [32] (2009) after 48 hours. However, one has to note that reductions in IHLC were comparable after 7% weight loss [32], supported by Haufe et al. [30] (2011) who also showed comparable reductions in IHLC after 6 months. Nevertheless, an increase in total energy expenditure by about ~50 kcal for every 10% decrease in the contribution of carbohydrates to total energy intake has been postulated [40], together with a decrease in ghrelin and leptin levels contributing to decreased appetite and satiety [41] following a LCD independently of body mass index (BMI). Importantly, these changes might be linked to an increase in ketogenesis and favorable changes in gut microbiota, which were even observed after an isocaloric LCD [42]. Another randomized controlled trial (RCT) aiming at maintained weight in adolescents reported a decrease in IHLC after 8 weeks only following an LCD (<25% C, 25% P, >50% F), but not a HCD (55% C, 25% P, 20% F) [43]. In summary, benefits from an LCD seem to include a favorable glucose metabolism (reduced insulin resistance [20], reduced basal glucose production [32]) independent of changes in IHLC, even in patients with established type-2 diabetes mellitus [44]. However, improvements of BMI, HDL and triglyceride profiles must be balanced with potential consequences of an LCD (i.e., high in dietary fat) such as elevated LDL and total cholesterol levels in the long-term [45,46]. Finally, both low carbohydrate consumption (<40% of total energy intake) and high carbohydrate consumption (>70%) were associated with higher overall mortality in unselected patients (i.e., a U-shaped relationship) [47], questioning long-term beneficial effects of LCD, but especially very-low-carbohydrate-diets (i.e., ketogenic diets).
Carbohydrate-insulin-model vs. energy-balance-model
Hypotheses discussing explanations for additional beneficial effects of an LCD on top of a hypocaloric diet be generated from the current discussion on two theories trying to explain energy metabolism in obesity: the carbohydrate-insulin-model and the energy-balance-model [48,49].
The carbohydrate-insulin-model focuses on the influence of dietary carbohydrates on the human body. Specifically, an increase in carbohydrates (i.e., high glycemic load) leads to increased insulin secretion (i.e., hyperinsulinemia) that promotes energy storage in adipose tissue, exacerbating hunger and lowering energy expenditure, all together promoting weight gain in a generally anabolic state [50]. By further stimulating glucose uptake, suppressing the release of fatty acids from adipose tissue, and promoting fat and glycogen production, hyperinsulinemia following carbohydrate intake induces a vicious cycle that “offers an explanation for why average BMI in many countries increased in the late 20th century as public health guidelines recommended replacement of dietary fat with carbohydrates, and consumption of high-glycemic-load foods increased substantially” [51]. Thus, the carbohydrate-insulin-model considers the high glycemic load as the starting point promoting anabolism including an anabolic hormonal profile, leading to “deposition” of substrates, leaving less energy for the brain (especially in the late postprandial period [52,53]) in turn inducing hunger and appetite [48].
Considering that insulin resistance is regarded a hallmark of NAFLD progression closely linked to inflammation, oxidative stress, and disease progression [54-56], an additional benefit of a LCD in NAFLD is reasonable from a pathophysiological perspective. Here, insulin resistance directly correlates with hepatic de-novo lipogenesis (DNL) [57], which has been shown to significantly contribute to IHLC in lean individuals without NAFLD (~11%), but being even more pronounced in obese individuals (~19%) and obese NAFLD patients (~38%). Most importantly, Luukkonen and colleagues [58] (2022) just recently described insulin resistance as an independent pathophysiological trait in NAFLD next to the genetic predisposition, being amplified if both factors are present. Considering this importance of insulin resistance in NAFLD, an increased DNL during carbohydrate overfeeding [59-61], an increased DNL in NAFLD [57,62], and the efficacy of LCD especially in hyper-insulinemic patients [40], LCD could offer a “way out” of this vicious cycle. Here, Cohen and colleagues [63] (2021) could already demonstrate a reduction of DNL within 8 weeks of dietary sugar restriction in adolescents.
In summary, specific beneficial aspects include the above-mentioned increase in energy expenditure [40,64], increase in satiety [41], lower insulin and ghrelin action in adipose tissue, higher glucagon action in non-adipose sites, and increased leptin sensitivity in the muscle [51].
The competing model to this theory is the energy-balancemodel that considers the increased availability of (cheap and energy-dense) food as the starting point for obesity [49]. Specifically, the brain regulates body weight in response to external signals from our food environment that stipulate hormonal signals controlling food intake, but also energy partitioning within the body [49]. Importantly, proponents of this model argue against the simplistic approach of the carbohydrate-insulin-model neglecting that several variables in the food environment influence energy intake and energy partitioning. For example, energy expenditure and energy intake are dynamically interrelated by physiological counteracting mechanisms (e.g., adaptive thermogenesis corresponding to a reduced energy expenditure if energy intake is decreased [65]) that are nearly impossible to look at in an isolated fashion [66]. While data supporting a lower energy expenditure following low-fat diets exist, authors claim that these differences are so small that “a calorie is a calorie” [66]. Also, one must acknowledge that evidence from meta-analysis is currently lacking that an LCD (favoring the carbohydrate-insulin-model) is more effective than a low-fat diet if calorie restriction is achieved (favoring the energy-balance-model) [37].
MEDITERRANEAN DIET (MD)
Looking beyond the macronutrient composition, it seems that the dietary composition is still relevant for the effect of a given diet on metabolic parameters. Here, a dietary composition according to the MD has been most consistently associated with improved phenotype of NAFLD [67]. Specifically, the MD has been defined “primarily a plant-based diet characterized by a high ratio of monounsaturated fatty acids (MUFA) to saturated fatty acids (SFA) with total fat accounting for 30–40% of daily energy consumption” [68].
Next to improvement in metabolic dysregulation [69] and prevention of cardiovascular diseases [70], adherence to the MD has been inversely associated with NAFLD prevalence [71] and severity [72-74], reduction in liver fat content [14,18,19,75-77], and LSM [18,78]. For instance, adherence to a low-carbohydrate MD (over 6 months) improved NAFLD (assessed by ultrasound) [77]. However, the inverse association between adherence to MD and decrease in liver fat content might be largely mediated (i.e., driven) by a decrease in BMI [74] emphasizing the central role of adipose tissue-liver crosstalk when studying liver-related outcomes [79].
Despite these promising results, the dietary composition of MD was heterogeneous across different studies and often combined with calorie restriction, thereby complicating direct comparison. Nevertheless, the best evidence that adherence to a MD on top of a hypocaloric diet is beneficial for NAFLD comes from studies from Israel. Gepner and colleagues [80,81] demonstrated that an LCD in combination with a MD achieved the greatest reduction in visceral adipose tissue and IHLC compared to an iso-caloric HCD. Interestingly, this effect was achieved despite only moderate weight loss, again supporting favorable effects of MD beyond calorie restriction [80]. Recently, the “DIRECT PLUS” RCT demonstrated a successful (and durable) weight loss and decrease in IHLC following a hypocaloric MD after 18 months [14]. What is even more interesting, the addition of dietary polyphenols (green tea and Mankai) further amplified these beneficial effects on IHLC (–38% relative change compared to –17% in the MD-only group) [14].
Specifically, several aspects seem to explain the success of the MD: First, one must consider that the MD is by itself characterized by a reduced carbohydrate intake (~approx. 40% of calorie intake), thereby mimicking favorable effects of a LCD on liver fat [82]. Second, the MD is low in food types that show clear harmful effects on NAFLD (such as Red and processed meat and SSB, as discussed in chapter Sugar sweetened beverages [SSB]), and rich in those that are considered beneficial (such as olive oil, nuts, legumes, seeds, whole grains, and vegetables) [67]. Third, the MD is rich in molecules/compounds that are generally regarded as “healthy”. Most prominently, polyphenols including flavonoids exhibit antioxidative effects reducing mortality in the general population [83,84], but also inhibit DNL, suppress the activation of hepatic stellate cells, and reduce carcinogenesis in animal models [85]. Carotenoids (i.e., lipid-soluble phytochemical) exert similar antioxidative properties [86] but are also discussed to decrease lipid accumulation, insulin resistance, oxidative stress, and inflammation in the liver [87]. Fourth, it still seems clear that the quality of ingested nutrients matters [36]. For example, 4 studies have shown favorable changes in IHLC if energy from fat is derived from MUFA and poly-unsaturated fatty acids (PUFA) compared to SFA following an isocaloric [88] or hypercaloric diet [89-91]. Also, an isocaloric diet high in MUFA was superior in reducing IHLC compared to isocaloric control diets despite unchanged body weight [92,93]. Finally, adherence to MD seems to be easier than to other diets (e.g., HCD), which has been demonstrated by the recent CORDIOPREV study reporting adherence to the MD in 7 of 8 patients over a period of 7 years, given that patients are supported by dieticians [94]. For the first time ever, a significant reduced incidence of major cardiovascular events in patients with coronary artery disease following a MD without energy restriction participating in this RCT was reported, further advocating this dietary composition [94].
FOOD GROUPS
Numerous food groups have repeatedly been associated with NAFLD [95]. Among them, red meat and SSB have shown the strongest negative impact on NAFLD prevalence and will be further discussed, while nuts and seeds seem to be protective [95,96].
Sugar sweetened beverages (SSB)
Dietary fructose intake—mostly via SSB and high-fructose corn syrup—is one of the food groups with the strongest evidence supporting harmful effects on multiple health outcomes, including NAFLD [97]. From a physiological point of view, fructose metabolization is nearly exclusively limited to hepatocytes [98]. By bypassing the rate-limiting step of glycolysis catalyzed by phosphofructokinase, fructose not only provides more substrate to DNL than glucose, but also occurs independent of insulin and the energy status of the cell [98], leading to an energy mismatch and subsequently promoting oxidative stress and insulin resistance [99,100]. Also, a roughly 100% first-pass effect following oral ingestion of fructose has been observed [101], suggesting metabolism in the liver directly upon consumption. Keeping this “fructose-processing burden” in mind, the harmful effect of significant and/or long-lasting fructose consumption on the liver seem reasonable.
In brief, several meta-analyses have tried to dissect the effect on glycemic control [102], metabolic syndrome [103], or NAFLD [104]. When fructose was substituted for other calories, no effect was evident regarding glycemic control compared. In contrast, a clearly harmful effect was observed when SSB were consumed on top of the usual diet (i.e., as excess calories) [102]: SSB showed a dose-dependent (increasing) effect on the prevalence of metabolic syndrome, while fruit juices showed a U-shaped relationship with protective effects at moderate doses [103]. Finally, a study on NAFLD found that addition of SSB (as ~30% excess energy) led to a significant increase in IHLC [104], while the beneficial effect when cutting down on fructose-containing sugars was less clear. However, all 3 available meta-analyses highlight the interaction with food sources (i.e., where excess fructose comes from) as an essential modifier of these effects, with SSB being the least favorable. Also, healthy individuals and/or adolescents seem to respond less to fructose supplementation [105] or restriction [106].
In individual studies, SSB have been associated with higher NAFLD prevalence [107-110], presence of NASH [111] and even a higher degree of fibrosis [112]. Recently, 4 RCT investigated the effect of fructose restriction on liver-related outcomes: Geidl-Flueck et al. [113] (2021) demonstrated a 2-fold increase in hepatic fatty acid-secretion rates in healthy men ingesting fructose/sucrose group vs. glucose sirup, Schwimmer et al. [114] (2019) reported a decrease in IHLC after 8 weeks of restricting free sugars, Simons et al. [115] (2021) showed a significant decrease in IHLC after 6 weeks of a fructose-restricted diet in NAFLD, and Khodami et al. [116] (2022) reported on an improvement of insulin resistance, steatosis, and fibrosis surrogates in NAFLD patients similarly restricting free sugars.
From a pathophysiological perspective, dietary fructose promotes DNL, impairs fatty acid oxidation, and triggers hepatic inflammation, thereby clearly fueling hepatic insulin resistance (reviewed in [99]). Also, epigenetic changes occur [117], and the role of the microbiome, metabolizing fructose to acetate being an additional substrate for DNL—is being increasingly understood [118]. Despite incompletely understood, dietary fructose even seems to increase nutrient absorption via improving survival of intestinal cells and increasing intestinal villus length [119].
Thus, although data regarding a long-term comparison between glucose and fructose consumption are lacking [36], available data clearly suggests that fructose consumption should be cut down to a minimum in patients with NAFLD.
Red and processed meat
Numerous studies within the last years have demonstrated a negative impact of red and especially processed meat on the prevalence of NAFLD. While some studies pointed towards a general association of meat with NAFLD [73,120,121], more recent observational longitudinal studies [122-124] and cross-sectional studies [125,126] have linked high consumption of only red meat to an increased prevalence of NAFLD [95]. Of note, white meat (i.e., chicken or turkey) did not show any significant associations [122], while processed meat of any type is still unfavourable [123,125]. Translating these associations into macronutrient composition, they are especially driven by animal protein since consumption of plant-based protein did not show a comparable association [121,127]. However, the harmful effects of high meat consumption on liver fat might be largely driven by a parallel increase in BMI [128], as also shown for the MD. Nevertheless, selected studies have even reported an increased risk of fibrosis in NAFLD patients with high red/processed meat consumption [123].
On a molecular basis, the diet-dependent acid-load seems to be an driving factor for these associations by inducing a low-grade metabolic acidosis129,130 leading to a disturbance in acid-base-homeostasis.131 Also, red meat contains a considerable amount of SFA and cholesterol, which have been shown to boost insulin resistance132 and drive hepatic lipid storage.133 Next, heme iron134,135 and nitrate (added for preservation) contribute considerably to the harmful effects of red or processed meat, potentially via increased oxidative stress.136 Finally, modification of the intestinal microbiota including the metabolism of certain components of red meat into harmful compounds (such as trimethylamine-N-oxide) seems to contribute to these negative effects.137,138 Focusing on processed meat, cooking meat at high temperatures for a long duration can form heterocyclic amines, which induce unfavorable health effects including an increased risk of cancer139 and chronic diseases, again mainly driven by an increased oxidative stress.140On a molecular basis, the diet-dependent acid-load seems to be an driving factor for these associations by inducing a low-grade metabolic acidosis [129,130] leading to a disturbance in acid-base-homeostasis [131]. Also, red meat contains a considerable amount of SFA and cholesterol, which have been shown to boost insulin resistance [132] and drive hepatic lipid storage [133]. Next, heme iron [134,135] and nitrate (added for preservation) contribute considerably to the harmful effects of red or processed meat, potentially via increased oxidative stress [136]. Finally, modification of the intestinal microbiota including the metabolism of certain components of red meat into harmful compounds (such as trimethylamine-N-oxide) seems to contribute to these negative effects [137,138]. Focusing on processed meat, cooking meat at high temperatures for a long duration can form heterocyclic amines, which induce unfavorable health effects including an increased risk of cancer [139] and chronic diseases, again mainly driven by an increased oxidative stress [140].
INTERMITTENT FASTING (IF)
Several types of “intermittent fasting” (IF) have gained increasing popularity in recent years. In brief, “time-restricted feeding” (TRF) involves calorie intake only during a pre-specified time window (usually for 4–10 hours). With regard to timing, a recent study applying TRF on healthy individuals indicates a certain benefit in glycemic control when feeding is restricted to the time between 06:00–15:00 vs. during the mid of the day (11:00–20:00) [141]. “Alternate day fasting” describes a mode of TRF in which fasting periods over 36 hours are followed by ad-libitum food consumption over the next 12 hours (i.e., every 2nd day, e.g., from 06:00–18:00). Finally, the 5:2 diet involves calorie restriction only on 2 non-consecutive days during the week, on which calorie intake is usually restricted to 500–600 kcal/day. This periodic calorie restriction is believed to provoke several physiological changes contributing to health benefits (reviewed in [142,143])—among others, it might counteract the disruption of circadian rhythm being associated with the development of NAFLD and metabolic syndrome [144,145].
Stimulated by the success of Stekovic et al. [146] (2019) demonstrating significant improvement of metabolic parameters after 4 weeks and 6 months, an increasing number of studies have elucidated the beneficial effects of IF on health outcomes. Lately, an umbrella review of meta-analyses of RCT studying obesity-related outcomes reported beneficial outcomes for BMI, body composition, serum lipids, glucose homeostasis, and blood pressure [147].
Focusing on NAFLD, 5 studies have so far specifically investigated IF in this patient population. Johari and colleagues [21] applied a modified alternate-day calorie restriction (i.e., 70% calorie restriction on fasting day, ad-libitum eating on non-fasting day) to demonstrate an improvement in ALT levels as well as LSM and ultrasound-based steatosis [21]. Another study showed a decrease in BMI and triglyceride levels following 12 weeks of ADF or time-restricted feeding (energy intake only during an 8 hours-window each day) despite no changes in LSM [148]. Holmer et al. [17] (2021) compared the 5:2 diet (<500/600 kcal/day on fast-days) with an LCD in patients with NAFLD. This diet was associated with a significant improvement in liver fat as assessed by MRI or CAP, as well as improvement in BMI and insulin resistance compared to a control diet, among others. However, no differences were observed compared to the LCD diet. Kord Varkaneh et al. [149] (2022) also compared the 5:2 diet over 12 weeks with a control group, and observed improvements of metabolic parameters including LSM and CAP. Finally, Xiao and colleagues [150] (2022) studied 60 NAFLD patients with type 2 diabetes mellitus randomized to 5:2 diet or liraglutide over 24 weeks, and found comparable metabolic improvement including a decrease in CAP in both groups. In addition to these studies, certain data exist on the effect of Ramadan fasting on the liver. Again, aside from the improvement in metabolic serum parameters including glucose homeostasis [151], non-invasive scores of fibrosis and markers of subclinical inflammation improved in NAFLD patients [152]. Also, Ramadan fasting reduced the gene expression of “fat-mass-and-obesity-associated protein” (FTO) in overweight/obese individuals [153], which has been associated with obesity [154] despite lower calorie intake [155].
However, it is currently a matter of debate whether IF (i.e., time-dependent calorie restriction) is more effective [156] or equally effective [157,158] than continuous calorie restriction (e.g., hypocaloric diet), and whether it is effective if no calorie restriction/dietary counselling is applied [159]. In the setting of type-2 diabetes mellitus [160], close monitoring of diabetes medication and blood glucose is needed due to concerns about hypoglycemia [161] although TRF has also been shown to be effective and safe in overweight/obese patients with type-2 diabetes mellitus. At the same time, sarcopenia might be an issue due to fasting inducing protein catabolism and muscle loss [162-164].
An often discussed effect of IF is an increase in ketogenesis (reviewed in [165]). In brief, the production of ketone bodies (mainly acetoacetate and β-hydroxybutyrate) from fatty acids serves as an alternative energy supply from the liver to peripheral tissues when carbohydrates are unavailable, therefore being pronounced during fasting or starvation [166]. At the same time, ketogenesis represents an alternative lipid disposal pathway metabolizing acetyl-CoA derived from β-oxidation. While NAFLD is characterized by an abundance of substrates that need to be metabolized by the liver inducing oxidative stress, DNL is upregulated [57,58,62,167] and ketogenesis downregulated, leading to an exhausted mitochondrial capacity [168]. Thus, on top of the direct beneficial effects of ketone bodies including antioxidative and anti-inflammatory functions (discussed in [169,170]), IF (but also very-low-carbohydrate-diets) could reverse this so-called “ketogenic insufficiency” that has been observed in NAFLD [171] by increasing hydrolysis of IHLC partitioning fatty acids towards ketogenesis, thereby improving mitochondrial redox state [20]. Additional beneficial effects of fasting might include the simulation of the peroxisome proliferator-activated receptor alpha (PPARα) /fibroblast growth factor 21 (FGF21) signaling [172] involved in regulating fatty acid metabolism [173].
PRECISION NUTRITION IN NAFLD
“Precision nutrition” aims at tailoring personalized dietary recommendations to individuals considering not only lifestyle and socioeconomic factors, but also incorporating data on the metabolome [174], microbiome and the genetic background [175]. Here, a huge effort is being made towards personalized medicine [176] and deeper understand the interactions between our diet and our environment. Although few studies have focused on patients with NAFLD, data from unselected cohorts focusing on clinical endpoints closely related to NAFLD are indeed astonishing. Here, Zeevi and colleagues [177] (2015) demonstrated that large interpersonal variability exists in the postprandial glycemic response to identical meals. Together with a follow-up study by their group again showing heterogenous glycemic responses to sourdough or white bread [178], these data indicate that often neglected factors such as the microbiome significantly influence the effectiveness of a given dietary intervention. Also, data from the PREDICT1 study support the central role of the gut microbiome explaining more variance in post-prandial triglyceride and insulin levels than the macronutrient composition of the ingested meals itself [179]. Exemplary looking at individual substrates, the beneficial effects of resveratrol on liver fat are discussed to be mediated by changes in the microbiome [180].
When trying to understand the influence of our genes on dietary responses, data show that they are highly relevant for our postprandial glucose response alone explaining ~50% of the variance [179]. Looking at individual single nucleotide polymorphisms (reviewed in [181]), the PNPLA3 rs738409 G-allele has been best studied as a modifier for the dietary response. An early study in Hispanic children indicated a significant positive correlation between IHLC and dietary carbohydrates only in homozygous carriers of the G-allele [182]. Also, following an LCD, the improvement in IHLC and insulin sensitivity was highest in G/G-carriers [183,184]. Similarly, two studies confirmed significantly larger changes in hepatic fat on a low n-6:n-3 PUFA ratio diet in homozygous carriers of the PNPLA3 risk allele [185,186]. Finally, dietary carbohydrates, but also polyphenols and PUFA were associated with significant fibrosis on histology only in carriers of the PNPLA3 G-allele (G/C or G/G) [187]. At the same time, our genes might not only influence our response to a certain diet, but also generally determine our macronutrient content. Here, it is believed that our genetic background explains up to 40% of our macronutrient intake [188] with SNPs in FGF21 (increased carbohydrate [189] or protein intake [190]) and FTO (increased protein intake [191,192]) being mostly studied, the latter potentially allowing greater weight loss during dietary/lifestyle interventions [193,194].
CONCLUSION
In summary, nutritional research und understanding the influence of diet on disease severity is one of the most complex aspects in the management of NAFLD patients. While being highly efficient when done consequently, evaluating the effects of dietary interventions is challenging as they impact on the whole metabolism, and specific (beneficial) effects on the liver are hard to detangle. While this makes firm conclusions and guideline recommendations difficult, this must not be misinterpreted as a limitation of dietary interventions per se. Currently, many roads seem to be leading to Rome as long as a calorie deficit is achieved and energy expenditure is increased. However, a hypocaloric diet, low in dietary carbohydrates, potentially including IF could be a diet tailored to successfully “treat” NAFLD, awaiting further study results. Also, increasing evidence suggests that a dietary composition according to the MD provides additional benefits for NAFLD patients beyond calorie restriction. On the other hand, personalized dietary recommendations might be necessary to make use of the full potential of dietary interventions in NAFLD.
Notes
Authors’ contribution
Georg Semmler: Conceptualization, Writing- Original draft, Writing- Reviewing and Editing. Christian Datz: Conceptualization, Writing- Original draft, Writing- Reviewing and Editing. Michael Trauner: Conceptualization, Writing- Original draft, Writing- Reviewing and Editing.
Conflicts of Interest
The authors have nothing to disclose regarding the work under consideration for publication. The following authors disclose conflicts of interests outside the submitted work: GS received travel support from Gilead. CD is part of the scientific advisory board of SPAR Österreich AG. MT received grant support from Albireo, Almylam, Cymabay, Falk, Gilead, Intercept, MSD, Takeda and Ultragenyx, honoraria for consulting from Albireo, Boehringer Ingelheim, BiomX, Falk, Genfit, Gilead, Hightide, Intercept, Janssen, MSD, Novartis, Phenex, Pliant, Regulus and Shire, speaker fees from Bristol- Myers Squibb, Falk, Gilead, Intercept and MSD, as well as travel support from AbbVie, Falk, Gilead, and Intercept. He is also co-inventor of patents on the medical use of 24-norursodeoxycholic acid.
Abbreviations
BMI
body mass index
CAP
controlled attenuation parameter
DNL
de-novo lipogenesis
HCD
high-carbohydrate diet
IF
intermittent fasting
IHLC
intrahepatic lipid content
LCD
low-carbohydrate diet
LSM
liver stiffness measurement
MD
Mediterranean diet
MUFA
mono-unsaturated fatty acids
NAFLD
non-alcoholic fatty liver disease
NASH
non-alcoholic steatohepatitis
PUFA
poly-unsaturated fatty acids
RCT
randomized controlled trial
SFA
saturated fatty acids
SSB
Sugar-sweetened beverages
TRF
time-restricted feeding